Algo de Antropología …

La Evolución del Hombre y la Gran Migración
elhistoriador.es/Open Mind(M.Martinón-Torres)/Investigación y ciencia(J.Osterkamp) — Los comienzos de la historia del hombre se sitúan en África. Allí se han encontrado los restos de los homínidos más próximos a él: los australopitecos, entre los que destacan el Australopithecus afarensis (entre 4 y 3 millones de años) y el Australopithecus africanus (entre 3 y 1,5 millones de años).
El primer hallazgo de estos seres fue realizado por Raymond Dart en el año 1924, en Taung (Transvaal). E1 escenario en el que apareció el hombre, el mismo que el de aquellos, es el inmenso territorio del África oriental, una sabana con masas arbóreas y numerosos lagos atravesado, desde el Mar Rojo al Transvaal, por la enorme falla llamada Valle del Rift.
El más antiguo representante de la especie humana es el Homo habilis, encontrado por primera vez por Louis Leakey en Olduwai (Tahzania) en los primeros años sesenta y del que se conocen algunos otros pocos ejemplares. Se le ha atribuido una antigüedad de 1,75 millones de años (es decir, que convivió con los australopitecos). Su capacidad craneana era de 750 cm3 y fue el autor de una industria de cantos talladas que se ha denominado Olduwayense (como los hallazgos de Melka Kunturé, en Etiopía, o Koobi Fora, al este del lago Turkana).
El siguiente homínido es el Homo erectus, cuyos primeros restos, por excepción, fueron encontrados por E. Dubois, en 1892, en los aluviones del río Solo (isla de Java), dándoseles el nombre de Pithecanthropus erectus, así como el Sinanthropus pekinensis, con varios ejemplares hallados en Chu-ku-Tien (cerca de Pekín) a partir de 1927. Ambos tipos se consideran como pertenecientes a Homo erectus en variedades alejadas por la geografía.
También lo son otros especímenes de África oriental, del norte de África (Ternifine, Argelia, y otros) y de Europa (mandíbula de Mauer, cerca de Heidelberg; Montmaurin, Alto Garona; Tautavel, Pirineos orientales, y otros). Los hallazgos europeos y sus yacimientos se situan entre 500.000 y 300.000 años, pero los africanos y los del oriente y sudeste de Asia tienen fechas atribuidas en torno al millón de años. La capacidad craneana de Homo erectus oscilaba entre 800 y 1250 cm3 y, desde sus orígenes en el barranco de Olduwai, produjo el Achelense, una industria carecterizada por los bifaces y los hendedores, con bolas y lascas.
En su haber cuenta la domesticación del fuego, lo que le permitió llegar a las regiones periglaciares de Eurasia y colonizar, a lo largo de más de un millón de años, todo el Viejo Mundo.

El Homo sapiens neanderthalensis apareció en Europa y en el Próximo Oriente hace unos 300.000 años, perdurando hasta las primeras fases de la última glaciación, la de Wur (hace unos 35.000 años). Se trató, casi con toda seguridad, de una evolución gradual y regional del Homo erectus.
Su voluminoso cráneo tenía una capacidad entre 1.300 y 1.750 cm3 y presentaba unos prominentes torus supraorbital y occipital. Fue el autor de un complejo de industrias unidas en el Musteriense y basadas en la producción de utensilios a partir de lascas (principalmente raederas). Además de los enterramientos ya citados, hay indicios de prácticas rituales: por ejemplo, el cráneo de un niño de unos doce años de Tashik Tash (Uzbekistán), estaba rodeado por un círculo de cinco pares de cornamentas de cabra salvaje.
Un hallazgo similar se produjo en la cueva Guattari (Monte Circeo, cerca de Roma): un cráneo de adulto fue vaciado por el occipital y rodeado de un círculo de piedras. Este caso sugiere el consumo ritual de la materia encefálica y, probablemente, lo atestigua de igual modo la antropofagia encontrada en la cueva de Krapina (Croacia). En sus fases finales, a causa del clima frío, el hombre de Neandertal utilizó ampliamente las entradas de las cuevas y los abrigos profundos (Abric Romaní, Capellades, Barcelona).
El Homo sapiens sapiens, el hombre moderno, tiene un origen muy antiguo que se sitúa fuera de Europa. Sus representantes más primitivos se han encontrado en África, donde tienen fechas de 100.000 años. Pero el exacto origen geográfico es desconocido.
En Europa apareció alrededor del 35.000. Se caracterizaba por la estabilización del volumen cerebral y la gracilización de su estructura ósea. Los diversos grupos de Homo sapiens sapiens han sufrido variaciones en función de la geografía y del clima de las regiones que progresivamente fueron ocupando, lo que explica las razas actuales.
En el África no mediterránea, con frecuencia se mantiene una situación prehistórica hasta tiempos recientes: en el desierto de Kalahari, África del Sur, subsisten aún pequeños clanes de cazadores recolectores en proceso de aculturación.
Por otra parte, el Homo sapiens sapiens llegó a Australia hace unos 40.000 años y, al igual que ocupó las regiones más frías del hemisferio septentrional, descubrió América o, en tiempos mucho mas recientes, gracias a una tecnología perfeccionada, alcanzó todas las islas del Pacífico.
Clasificación y Evolución de los Homínidos

La clasificación de los seres humanos como uno más de los animales que pueblan la Tierra se establece siguiendo las mismas pautas que para éstos: el hombre pertenece al tipo cordados, clase mamíferos, orden primates, suborden catirrinos, infraorden antropomorfos, familia homínidos, en la cual se incluyen el género Homo y la especie sapiens. Los más de dos mil restos fósiles humanos hallados hasta el momento han hecho que las denominaciones aludan, unas veces, al origen geográfico del yacimiento (hombre de Neandertal, por la localidad alemana de este nombre) y otras, a la posición que ocupa dentro del árbol genealógico de la especie humana.



Los primeros homínidos aparecieron a finales de la era terciaria, hace aproximadamente unos quince millones de años. Como miembro del orden de los primates, el ser humano comparte hasta un 99 % de macromoléculas con los otros individuos del mismo orden; por tanto, debe de existir un antepasado común entre nuestra especie y los Australopithecus, el género más parecido al Homo.
Los australopitecos, o monos meridionales, han sido localizados en África meridional y oriental, y su cronología abarcaría de los 6,5 millones de años de los Preaustralopithecus a un millón de años cuando se extinguieron. A lo largo de este largo período pueden subclasificarse en varios grupos y subgrupos, que se diferencian por sus características morfológicas.
¿Cuántos seres humanos ha habido?

Cien mil millones de seres humanos nos han precedido [La coronación de la Virgen, o El paraíso, de Tintoretto, primer boceto, Museo del Louvre].
Los escolásticos medievales ya intentaron dar con una respuesta, por preocupaciones de índole práctica: ¿cuántos caben en el cielo? El infierno, ¿se llenará algún día? ¿Cuántos son los elegidos que al final del mundo irán al cielo? Al menos esto último lo respondía su fuente primaria, y con una cifra tan baja que da escalofríos: 144.000 (Apocalipsis 14: 1, 3). Pero 144.000, ¿entre cuántos más?
Una fórmula para calcular el tamaño de la humanidad
Fue el influyente demógrafo canadiense Nathan Keyfitz, fallecido hace unos años, quien abordó matemáticamente la pregunta de cuál es el número de los que han vivido. Su receta es una fórmula en la que entran todas las magnitudes que influyen en la contribución positiva al crecimiento del número de seres humanos; el parámetro principal es la tasa de natalidad humana, que depende de la esperanza de vida de cada generación y del tiempo en que estén activas reproductivamente.
Partiendo del tamaño mínimo concebible de la población (dos, uno de cada sexo) en un instante de tiempo x (sobre esto volveremos más adelante), se llega, con todos esos parámetros y la fórmula, a una curva ascendente de la población de x en adelante.
Se pueden entonces integrar matemáticamente las gráficas resultantes para obtener el número de todas las personas que han ido viviendo en cualquier corte temporal. Ni que decir tiene, el diablo está en los detalles que salen al paso en la fórmula de Keyfitz. Los amigos de las matemáticas pueden encontrar en otras partes una explicación de la fórmula y su deducción.
Aquí cabe dar en forma de tabla cuántas personas vivían en distintos momentos y su suma según la Oficina de Referencia sobre la Población, una organización estadounidense:
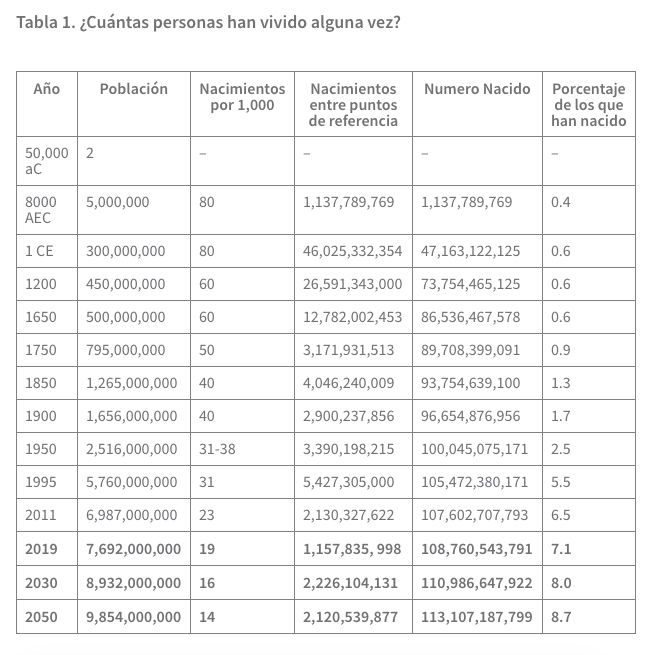
Pese a todas las matemáticas exactas, el conjunto no deja de ser una cábala bastante arriesgada, ya que se deben hacer diversas suposiciones sin mucho fundamento. Así, los investigadores intuyen que la población humana de la Tierra no ha ido creciendo siempre de forma paulatina y sin interrupciones, conforme a una curva ascendente expresable matemáticamente de manera sencilla, sino que, según las épocas, unas veces ha aumentado más deprisa y otras más despacio.
Y con bastante seguridad, hasta ha disminuido en ocasiones, incluso de forma drástica: por catástrofes naturales o por otras razones, ha tenido que pasar por «cuellos de botella» demográficos.
Es conocida, por ejemplo, la polémica entre los investigadores sobre la erupción del supervolcán Toba, de hace 74.000 años, cuyas consecuencias mundiales debieron de afectar gravemente a la joven humanidad.
Como, por lo tanto, no se puede tomar como premisa un crecimiento continuo de la población humana desde los primeros Homo sapiens (de hace al menos, quizá, unos 300.000 años), los calculadores de la población inteligentes recurren a un truco: sitúan ese punto x, en el que vivieron los dos seres humanos que empezaron la producción de descendencia, en un instante lo más «moderno posible»; eligen, en concreto, que el origen fuese 50.000 años antes del principio de nuestra era.
Con ello se decreta que la mayor parte del tiempo en que ha habido Homo sapiens (parece seguro que los primeros seres humanos modernos vivieron mucho antes) es irrelevante, ya que no afecta demasiado al resultado final.
Catástrofes y crecimiento vacilante de la población
La población creció al principio muy lentamente: según las fórmulas de Keyfitz y ese punto de partida escogido, entre el 50.000 y el 8000 antes de Cristo (momento en que se calcula que vivían unos cinco millones de personas) nacieron y fallecieron solo mil millones y pico de seres humanos. No muchos más habría que sumar si el punto de partida x se situase más en el pasado (al menos, si la pizca de unos cuantos millones de personas no importa mucho).
La mayoría de los paleodemógrafos coincide, en cualquier caso, en que durante toda la Edad de Piedra fueron pocos los seres humanos que habitaron la Tierra. Pero sobre todo es seguro que esos pocos seres humanos vivían poco (la esperanza de vida al nacer era de diez años).
A lo largo de los cientos de miles de años anteriores al principio de nuestra era, en los que se inventó, entre otras cosas, la agricultura, el aumento de la población se aceleró mucho: más y más muertos y vivos hincharon el número total de los seres humanos.
Los demógrafos han podido evaluar las tasas de natalidad probables de las épocas siguientes cada vez mejor (aunque no precisamente más allá de toda duda). Piensan que al principio de nuestra era (cuando vivían unos 300 millones de personas) era un asombroso 80 por 1000 (la tasa de natalidad siempre se da en nacidos al año por cada mil personas de la población total); para 1750 había bajado a 50. Como comparación: la tasa de natalidad de Alemania era en 2017 de solo 8,6, y la de España, de 9,2; las tasas de natalidad más altas, las de algunos países africanos, no llegan actualmente a 50, y el promedio es de alrededor de 18.
Por lo tanto, la tasa de natalidad se ha hundido en nuestros tiempos, pese a una esperanza de vida y un período de reproducibilidad mayores y pese a los grandes avances médicos. Pero como la población es mucho mayor que en la Edad de Piedra, la cifra total de los seres humanos que ha habido crece hoy mucho más deprisa que antes. Y por lo tanto hay que hacer una advertencia: las respuestas a nuestra pregunta, por lo demás vagamente calculables, se quedan desfasadas cada vez más deprisa.
Antropología: qué hemos aprendido en la última década

El uso cotidiano de la palabra investigar está henchido de connotaciones sobre progreso y futuro. Paradójicamente, el origen etimológico de este verbo está en el vocablo latín investigare, que viene a su vez de la palabra vestigium (vestigio, huella).
Así, el significado original del término investigar sería el de «ir tras la huella». Para aquellos que estudiamos periodos antiguos, volver a la raíz de la palabra investigar pone de manifiesto una noción con frecuencia olvidada y es que, para poder progresar, es necesario conocer y aprender de lo pasado. La paleoantropología es la disciplina que estudia el origen y la evolución del hombre y trata de reconstruir la historia de los cambios biológicos y culturales experimentados por nuestros ancestros desde que, hace unos seis millones de años, se separaron los linajes que dieron lugar a humanos y chimpancés.
Uno de los principales cuerpos de evidencia sobre los que se sustentan los estudios sobre evolución humana son los fósiles de especies de homínidos extintas. Esto conlleva, con frecuencia, a la idea errónea de que la paleoantropología es un ámbito de estudio encapsulado en el pasado, con contribuciones mínimas, como mucho anecdóticas, a nuestra comprensión del ser humano actual.
Justamente, la investigación desarrollada en evolución humana durante la última década ha roto este paradigma, tanto desde el punto de vista metodológico como conceptual, desarrollando investigaciones en la frontera del conocimiento y aportando datos inéditos sobre nuestra propia especie.
El pasado se descifra ahora con la tecnología del futuro. La necesidad de hacer «hablar a los muertos», y de maximizar la información que se puede extraer del preciado y escaso registro fósil y arqueológico, ha llevado a los paleontólogos y arqueólogos a perfeccionar y exprimir las aplicaciones de los métodos actuales, delineando, en algunos casos, nuevas líneas de investigación. La aplicación, por ejemplo, de técnicas de imagen de alta resolución al estudio de los fósiles, ha permitido la fundación de una rama metodológica, con entidad propia, conocida como Antropología Virtual.
Así, la era digital llega también al mundo del pasado y, con estas técnicas, es posible ahora estudiar y manipular de forma no destructiva un fósil, realizando mediciones y reconstrucciones bidimensionales y tridimensionales (2D, 3D) de cualquier superficie tanto externa como interna del objeto de estudio. Pero sin duda, el mejor exponente de la fructífera relación entre la tecnología y el estudio del pasado se da en la consolidación de un ámbito de investigación reciente, el de la paleogenética o análisis de ADN antiguo.
El reconocimiento con el Premio Princesa de Asturias de Investigación Científica y Técnica 2018 al biólogo sueco Svante Pääbo, considerado uno de los fundadores de la paleogenética, es sintomático de la trascendencia que han tenido los estudios moleculares en la reconstrucción de una parte fundamental de nuestra historia. El equipo liderado por Svante Pääbo, director del Instituto Max Planck de Antropología Evolutiva de Leipzig (Alemania), ha sido pionero en la adaptación de técnicas de alto rendimiento de secuenciación del ADN al estudio del ADN antiguo, posibilitando así el análisis del genoma completo de organismos extinguidos.
Precisamente, su colaboración con el equipo investigador de los yacimientos de la sierra de Atapuerca (Burgos), ha posibilitado el hito científico de extraer ADN de los homininos del yacimiento de la Sima de los Huesos (430.000 años de antigüedad), el ADN más antiguo jamás recuperado hasta el momento en un ambiente sin permafrost.

Entrada principal a la Gran Cueva de Niah, la cueva de piedra caliza más grande del mundo, que se encuentra en el Parque Nacional Niah, en el estado malasio de Sarawak, en la isla de Borneo.
Las cuevas de Niag contienen los restos más antiguos de Homo sapiens encontrados en Borneo.
Estudios publicados recientemente demuestran que los indicios de actividad humana en estas cuevas se remontan a entre 46.000 y 34.000 años
La consolidación de los estudios de paleogenética ha proporcionado datos sobre el origen de nuestra especie, Homo sapiens, y el tipo de interacción que mantuvimos con otras especies de homininos ya extinguidos, que parecían impensables hace una década.
Hasta ahora, la historia del origen de Homo sapiens cabía en un relato casi lineal sobre el surgimiento en un lugar único de África.
Apoyándose en un abanico de capacidades físicas e intelectuales avanzadas, los humanos modernos se habrían expandido por todos los continentes no hace más de 50.000 años. Este es el cuerpo de la conocida como teoría del Out of Africa, en la que se sugiere que en su expansión por todo el planeta, Homo sapiens habría reemplazado a todos los grupos humanos arcaicos sin cruzarse genéticamente con ellos.
Sin embargo, los análisis moleculares han roto este paradigma con la revelación de que los humanos modernos no solo hibridaron y tuvieron descendencia fértil con especies humanas extintas, como los neandertales, sino que entre el 2% y el 4% de los genes de las poblaciones no africanas actuales pertenecen a estos. Pero esto no es todo.
A partir del análisis genético de un hueso de la mano, hallado en la cueva de Denisova, en los montes Altai (Siberia), los genetistas han identificado una nueva población humana, de la que apenas tenemos fósiles y por lo tanto desconocemos su apariencia física, pero que es genéticamente diferente de neandertales y humanos modernos.
Esta población, coloquialmente conocida como los denisovanos, habría coexistido e hibridado con nuestra especie y con H. neanderthalensis de forma que, entre el 4% y el 6% del material genético de los humanos actuales que pueblan Papúa Nueva Guinea, Australia y Melanesia, pertenecería a los denisovanos.
La paleogenética se erige como una investigación de frontera en muchos de los sentidos descritos por el filósofo de la ciencia Thomas Khun, en su obra La estructura de las revoluciones científicas, en cuanto ha permitido utilizar metodologías (la genética) y conceptos (hibridación) que no eran comunes en el ámbito paleoantropológico, proporcionar resultados inesperados que ponen en cuestión el paradigma imperante y, sobre todo, abrir puertas a dimensiones imprevistas del conocimiento.
Si bien en la actualidad somos la única especie humana sobre la faz de la tierra, ahora sabemos que no siempre estuvimos solos, y que conocimos y tuvimos descendencia con otros humanos que ya no existen pero que han dejado su firma en nuestros genes.
El vértigo intelectual se desata: ¿Qué significa ser híbrido? ¿Cómo fue conocer y convivir con otra especie humana inteligente? ¿Qué implica tener en nuestra sangre material genético de otras especies humanas? ¿Tuvo que ver esta hibridación en la extinción de nuestros hermanos los neandertales? ¿Aporta esto una perspectiva diferente sobre la diversidad humana actual?
Así pues, aunque es difícil sustraerse a la fascinación que ejerce el conocimiento sobre los periodos más antiguos, de los que la evidencia disponible es todavía más escasa, la investigación en el ámbito de la paleoantropología en la última década ha sido particularmente prolija en información sobre el periodo más reciente y, en especial, sobre la historia de nuestra propia especie.

Cueva desinova
Los datos genéticos, junto con nuevos descubrimientos fósiles y nuevas dataciones que sitúan a nuestra especie fuera del continente africano antes de lo estimado por la teoría del Out of Africa, han supuesto la revelación de un pasado completamente desconocido para las poblaciones actuales.
En este marco, el presente artículo repasará algunos de los hitos principales descubiertos en esta última década en el ámbito de la evolución humana, pero con un énfasis particular en la idea de que, como expresó el filósofo Martin Heidegger «la forma suprema del saber es la pregunta» y el diagnóstico del estado de salud de una ciencia vendrá dado, ya no tanto por su capacidad de generar respuestas, sino de generar preguntas nuevas, en este caso, sobre nosotros mismos.
NUESTRO ORIGEN HÍBRIDO
Uno de los principales problemas que plantea el tema de las especies híbridas es que atenta contra el concepto biológico de especie originalmente planteado por el biólogo evolutivo Ernst Mayr. Según Mayr, una especie es un grupo o población natural de individuos que pueden cruzarse entre sí, pero están aislados reproductivamente de otros grupos afines.
Este concepto implica que individuos de especies diferentes no deberían de poder hibridar ni tener descendencia fértil con otros individuos que no pertenezcan al mismo taxón. Sin embargo, la naturaleza nos proporciona un abanico amplio de casos de hibridación, principalmente en el mundo vegetal, en las que las especies de ascendencia mixta pueden tener hijos que no son necesariamente estériles.
En el mundo animal son menos frecuentes o menos conocidos, aunque se conocen casos entre especies de perros, gatos, osos, ratones y primates, como los monos aulladores. Un ejemplo particularmente interesante del mundo de los primates es el de los híbridos que resultan del cruce entre diferentes especies de monos del género Cercopithecidae, conocidos familiarmente como babuinos o papiones y que, como veremos, nos han proporcionado información muy útil.
Otro problema adicional es que, hasta la aplicación de las técnicas de estudio molecular al registro paleontológico, el concepto de especie biológica era difícilmente constatable en el registro fósil. Salvo que dispusiésemos de una máquina que nos permitiese viajar en el tiempo, resulta imposible comprobar si individuos de especies diferentes que se han extinguido pueden (y han) interaccionado sexualmente y si de estos cruces se ha obtenido progenie fértil.
Por otra parte, existe muy poca información sobre cuál es la apariencia física de los híbridos y de ahí su dificultad para reconocerlos en el registro fósil.
En el ámbito paleontológico, el concepto de especie utilizado es mucho más pragmático, y este término suele utilizarse como categoría útil en la que agrupamos a aquellos individuos que consideramos que, principalmente por sus características anatómicas (aunque idealmente debiéramos incluir su comportamiento y nicho ecológico), forman un grupo homogéneo y en principio reconocible y distinguible de los individuos de otros grupos.
Esta distinción morfológica clara entre los individuos que potencialmente asignamos a una especie, se utiliza como indicador indirecto del aislamiento que ha habido entre esta y otras poblaciones (especies) que, de haberse cruzado de forma sistemática, no habrían conservado su morfología característica. ¿Invalida el hecho de que se hayan producido cruces entre Homo sapiens y Homo neanderthalensis el hecho de que son especies diferentes? No necesariamente, aunque el debate está servido.
Los neandertales son una especie de hominino cuyo origen se remonta a las poblaciones que habitaron Europa hace unos 500.000 años y cuya extinción tuvo lugar hace unos 40.000 años coincidiendo en términos generales con la entrada de los humanos modernos en Europa, razón por la que se plantea que nuestra especie haya jugado algún papel en su extinción.

Homo neanderthalensis
Pocas especies del género Homo han levantado más pasiones que los neandertales dada su proximidad a nuestra especie y su fatídico destino. El estudio de los restos esqueléticos y arqueológicos atribuidos a esta especie permite retratar a un humano con un cerebro de igual tamaño o incluso ligeramente superior al nuestro, con demostradas capacidades intelectuales y físicas, grandes cazadores, pero también expertos conocedores de la utilización de recursos vegetales, no solo como alimento sino por sus propiedades medicinales.
Los neandertales también enterraban a sus muertos y utilizaban ornamentos y si bien su expresión simbólica y artística parece menos explosiva que la de los humanos modernos, no difería sustancialmente de los Homo sapiens de su tiempo. Un error común en el sempiterno debate sobre la posible «superioridad» cultural de Homo sapiens frente al Homo neanderthalensis es el anacronismo de comparar la producción artística de estos últimos con la de los humanos modernos actuales, en vez de con los grupos de los que fueron contemporáneos.
La reciente datación de pinturas rupestres de varias cuevas españolas en Cantabria, Extremadura y Andalucía, arrojan fechas anteriores a la llegada de los humanos modernos a Europa, dejando así abierta la posibilidad de que hayan sido los neandertales los autores de este tipo de manifestación artística.
¿Siendo por lo tanto tan sofisticados y tan parecidos a nosotros en cuanto a su comportamiento complejo, y sabiendo que ha habido cruces genéticos entre ambas poblaciones, podemos considerarlos especies diferentes?
A este debate se suma la noción de que, según estudios genéticos publicados a finales del año 2018 por Viviane Slon y sus colegas, los episodios de hibridación entre neandertales, humanos modernos y denisovanos, habrían sido relativamente frecuentes, lo que en principio cuestiona la premisa de la existencia de una barrera (biológica, cultural o geográfica) necesaria para el aislamiento reproductivo entre dos especies supuestamente diferentes.

Un investigador del laboratorio de la Universitat Autónoma de Barcelona prepara un espécimen prehistórico para testar su contenido de ADN.
Este espécimen se desenterró en un yacimiento prehistórico de la isla de Menorca, España
Los datos fósiles y genéticos disponibles en la actualidad, todavía no son suficientes para poder llegar a una conclusión clara.
Sin embargo, es importante resaltar que aun cuando hablemos de cruces relativamente frecuentes entre individuos de diferentes especies (algo sobre lo que la naturaleza ya nos ha proporcionado ejemplos en especies vivas), esto no significa necesariamente que los cruces sean la norma en la historia biológica y demográfica de ese grupo.
Existen determinadas circunstancias que pueden hacer que esos cruces sean más propicios, como periodos en los que una población sufre algún tipo de debilidad demográfica, una porción de esa población, quizá marginal, que encuentra dificultades para procrear dentro de su propio grupo o, principalmente, en zonas de transición ecológica, a caballo entre dos ecosistemas diferentes en los que encontrábamos dos especies con adaptaciones distintas.
El propio Ernst Mayr matizó que los mecanismos de aislamiento por los que un linaje estaba separado reproductivamente de otro se referían a las propiedades biológicas de los individuos que prevenían el entrecruzamiento habitual de estas poblaciones, admitiendo que aunque se podían producir cruces ocasionales, el carácter de este intercambio no tenía la entidad suficiente como para considerarse que se había producido la fusión completa entre las dos especies.
Y es ahí donde radica el quid de la cuestión. Los neandertales son, probablemente, una de las especies humanas mejor caracterizadas y conocidas de todo el registro fósil.
El cráneo bajo y alargado con una protuberancia obvia en su parte posterior («moño occipital»), la proyección característica de su cara alrededor de la región nasal (lo que técnicamente se denomina prognatismo medio-facial) y su marcado reborde óseo en la región de las cejas (toro supraorbitario), son algunas de las características neandertales que, desde su origen hace casi medio millón de años en Europa hasta su extinción, se mantuvieron virtualmente intactas.
Si se hubiera producido una mezcla sistemática y habitual entre neandertales y humanos modernos, cabría esperar un «atenuamiento» o modificación de estos patrones. Sin embargo, son precisamente los neandertales tardíos los que presentan las morfologías más típicas y pronunciadas. A esto se añade que, salvo por los datos proporcionados por los análisis genéticos, carecemos de yacimientos en los que se pueda constatar la convivencia de ambos grupos.
Los yacimientos del Monte Carmelo, en Israel, representan el mejor testimonio de la proximidad física entre ambas especies, pero en todos los casos la evidencia de la presencia de uno u otro grupo aparece siempre en niveles intercalados, nunca en el mismo. En contra de lo que reza la expresión popular, neandertales y humanos modernos habrían estado «revueltos, pero no juntos», y aunque se hubieran podido dar cruces ocasionales, esta no fue la norma, ni el hombre actual puede considerarse una fusión de ambos. En este sentido, la hipótesis de que los neandertales se extinguieron porque fueron absorbidos por los humanos modernos perdería fuerza.

Ejemplar de hamadriade (Papio hamadryas), del género Cercopithecadae. Los estudio realizados a especímenes resultantes del cruce entre diferentes especies de esta familia han proporcionado información muy útil para el estudio de la hibridación de homínidos
Una última apreciación interesante sobre los híbridos entre especies viene de los estudios que investigadores como Rebecca Ackerman y su equipo han realizado en monos babuinos.
Es una idea establecida en el imaginario popular, que un híbrido tiene que presentar una morfología intermedia entre las especies parentales, o bien un mosaico, una combinación de rasgos de una y de otra.
Sin embargo, Ackerman y sus colegas revelan que, con frecuencia, los híbridos no se parecen «ni a su padre ni a su madre», y en muchos caracteres se producen «novedades morfológicas». Los híbridos suelen presentar un tamaño claramente mayor o menor que sus padres y un número elevado de patologías y anomalías poco frecuentes en las poblaciones originales.
Algunas de estas anomalías, como alteraciones en las suturas cráneo-faciales, patologías dentales bilaterales o asimetrías craneales, son sin duda un reflejo de «desajustes» en el desarrollo. Así, aun cuando la hibridación sea posible, el encaje estaría lejos de ser perfecto.
Precisamente en el ámbito de la paleogenómica, existen estudios como los de Fernando Méndez y su equipo en los que se plantea la posibilidad de que los humanos modernos hubieran desarrollado un tipo de respuesta inmune contra el cromosoma Y neandertal, de forma que los embarazos de fetos masculinos híbridos, hijos de madre sapiens y padre neandertal, terminarían de forma recurrente en abortos naturales.
De esta forma, y al tiempo, la preservación de la información genética neandertal se vería amenazada.
En esta línea, podríamos interpretar que si bien la hibridación no fue la causa de la extinción de los neandertales pudo ser un factor coadyuvante en la diezma de esta especie. Lejos del clásico ejemplo de los dinosaurios y la caída de meteoritos, las extinciones en el mundo animal son generalmente procesos lentos, delicados juegos de equilibrio demográfico en los que no hacen falta grandes eventos o catástrofes para que las balanzas se inclinen hacia un lado u otro.
Mientras la fertilidad de los neandertales podría haberse visto debilitada con la mezcla, algunos expertos sugieren que de la hibridación con neandertales y denisovanos nuestra especie habría adquirido secuencias genéticas ventajosas para la adaptación a nuevos ambientes cuando abandonamos África, principalmente a través de cambios en el sistema inmune. Así, la hibridación podría haber sido ventajosa para nuestra especie como fuente de genes beneficiosos en nuestra conquista del mundo.

Pintura de la Cueva de Altamira, en Cantabria, España, que data del Paleolítico Superior.
Es necesario seguir investigando para poder ahondar el efecto que el intercambio genético ha tenido en el porvenir de cada una de estas especies.
Sin embargo, y aun cuando la hibridación haya podido afectar negativamente a denisovanos y/o neandertales, puesto que son estos los que se han extinguido, la paleogenética sugiere que la interacción de los humanos modernos con las especies extintas no fue necesariamente violenta.
Una de las teorías más clásicas que se han planteado para explicar la extinción de los neandertales es la de la confrontación, incluso violenta, entre ambas especies.
Homo sapiens ha sido descrita como una especie altamente «invasiva» y su paso, como el caballo de Atila, se ha asociado a la extinción de muchas especies de grandes animales (megafaunal extinction), incluyendo a los neandertales. Aunque sexo no implique necesariamente amor, el hecho de que conservemos un porcentaje de ADN neandertal en nuestras venas sugiere que alguien se tuvo que preocupar de criar y asegurar la supervivencia de los hijos híbridos, una realidad que quizá nos permite suavizar el estereotipo de un sapiens violento y avasallador.
LA EVIDENCIA «DURA» DE NUESTRO ORIGEN
No cabe duda de que el avance tecnológico de los últimos años nos lleva al examen de lo pequeño y lo molecular. A la revolución metodológica del análisis genético se suma el nacimiento de la paleoproteómica (estudio de las proteínas antiguas), una disciplina que dará mucho que hablar en la próxima década.
Sin embargo, el núcleo duro de la antropología, el corazón, ha sido y lo seguirán siendo los fósiles. Sin fósiles no habrá ADN, ni proteínas, faltará la fuente de evidencia más duradera y más completa de la que se alimenta la paleoantropología.
Surgen técnicas de análisis casi futuristas y sin embargo la realidad nos recuerda, literal y figuradamente, que, para poder avanzar en este campo es necesario «tener los pies en la tierra», esa tierra que hay que excavar y de la que saldrán, a golpe de trabajo y caprichosa suerte, los huesos de nuestros antepasados. Falta todavía mucho terreno por conocer.
Existen grandes vacíos de fósiles en los mapas, regiones como el oeste africano, el subcontinente indio o la península arábiga de los que apenas se dispone de registro, de forma que los hallazgos en estas zonas son como piezas nuevas que nos obligan a rearmar el puzle.
En la última década, antiguos y nuevos fósiles encontrados en Asia están desplazando el epicentro de atención al continente asiático y cabe augurar muchas sorpresas. Incluso en la reconstrucción de la historia de nuestra especie, contada hasta ahora en clave exclusivamente africana, los fósiles hallados en esta última década tienen algo que decir.
Desde el punto de vista fósil, la hipótesis del origen africano de nuestra especie se ha sustentado principalmente en el hallazgo en África de los restos más antiguos que pueden atribuirse a Homo sapiens.
Entre estos destacan los cráneos de Herto y Omo en Etiopía, datados entre 160 y 180.000 años de antigüedad. Además, en el Oriente Próximo, los yacimientos de Qafzeh y Skhul han proporcionado una colección importante de fósiles también atribuidos a nuestra especie que datan entre 90 y 120.000 años.
Sin embargo, según el Out of Africa, no era hasta hace unos 50.000 años, cuando nuestra especie era capaz de adentrarse en Europa y Asia por lo que a la presencia de Homo sapiens en Israel no se le daba la categoría de dispersión o «salida» propiamente dicha.

Sin embargo, en la última década, un número importante de fósiles cuestionan la fecha de los 50.000. Entre esos fósiles cabe destacar los dientes y mandíbula hallados en Daoxian (Fuyan Cave) y Zhirendong, en el sur de China, o la falange de la mano recuperada en Al-Wusta (Arabia) que sitúan a nuestra especie fuera de África al menos hace 80.000 años, aunque su presencia podría incluso adelantarse a los 100.000. En la misma línea está el hallazgo de un maxilar humano en el yacimiento de Misliya (Israel) que, con una edad en torno a los 190.000 años, es tan antiguo como los fósiles más antiguos africanos atribuibles a Homo sapiens.
Cada vez es más patente que Homo sapiens fue capaz de adaptarse a otros territorios antes de lo pensado, si bien este sigue siendo un debate abierto. La evidencia genética, aunque discutida, sigue apuntando a que la humanidad actual proviene principalmente de una dispersión sucedida hace unos 50.000 años. Sin embargo, esto no significa que no hubiera habido otras salidas anteriores de las que quizá no haya quedado huella en los humanos modernos de hoy o quizá no hemos sido capaces de detectar todavía.
Constreñidos por la utilización de esquemas con flechas para representar las dispersiones de los humanos en un mapa, olvidamos que las migraciones de los homininos no son movimientos lineales y direccionales, con un propósito o meta, a modo de marcha o excursión. Las migraciones humanas, como las de cualquier otro animal, deberían representarse como expansiones o ensanchamientos del rango de ocupación de un grupo que, ante la falta de barreras (ecológicas o climáticas, por ejemplo), y en condiciones demográficas favorables, aumenta su territorio de asentamiento.
Es probable que el Out of Africa no fuese un único evento o viaje sino un flujo más o menos continuo, de caudal variable. Quizá no hubo uno, sino muchos Out of Africa, pero también varios Into Africa, reentradas que no son técnicamente reentradas, porque los homininos «no vuelven» como quien vuelve a su hogar, sino que se expanden el diámetro de su territorio siempre que no haya algo que se lo impida.

Cráneo de un adulto humano con las vértebras del cuello, de la Sima de los Huesos, del yacimiento de Atapuerca, en Burgos, España, descubierto en 1984.
Los dientes, la mandíbula y los huesos faciales parecen neandertales, mientras que el cráneo es primitivo.
Esto sugiere que los rasgos característicos de los neandertales evolucionaron de manera separada, un modelo evolutivo conocido como modelo de acreción.
Por último, los hallazgos fósiles del yacimiento de Jebel Irhoud, en Marruecos, con 300.000 años de antigüedad, arrojan nueva luz (¿o más interrogantes?) sobre el origen de nuestra especie.
Aunque estos especímenes carecen de algunos de los rasgos que se consideran exclusivos de Homo sapiens (como la barbilla, la frente vertical o el cráneo alto y abombado), muchos investigadores los consideran los representantes más antiguos de nuestro linaje.
La novedad radica ya no tanto en su antigüedad como en su localización. La mayoría de los fósiles africanos atribuidos a nuestra especie se habían registrado, hasta ahora, en regiones del este o del sur de África, pero no del norte.
A raíz de este y otros descubrimientos fósiles en África, está cobrando peso la hipótesis de que nuestro origen no está en una sino en varias poblaciones que llegaron a habitar en regiones muy dispares del extenso continente africano y mantendrían intercambios genéticos intermitentes. Homo sapiens habría evolucionado en un patrón más reticulado y menos lineal de lo que se había creído hasta ahora. Esta teoría comienza a conocerse como el «Multirregionalismo africano» y apunta a que, en sus raíces más profundas, Homo sapiens era ya una mezcla, un crisol de poblaciones diversas, con una gran variabilidad física y cultural.
Curiosamente, el término «multirregionalismo» hace referencia a otra de las grandes teorías que durante el siglo XX se ha esgrimido para explicar el origen de Homo sapiens. El «Multirregionalismo», en contra del Out of Africa, mantenía que el origen de nuestra especie estaba en la evolución paralela de varios linajes que, en diferentes partes del mundo, daban lugar a Homo sapiens.
El multirregionalismo también observaba el intercambio genético entre estos grupos paralelos. Sorprendentemente, quién nos lo hubiera dicho, esta década ha sido testigo de cómo dos polos opuestos (Out of Africa y Multirregionalismo) han aproximado posiciones.

Un trabajador del Instituto Max Planck taladra un hueso de neandertal para obtener su ADN
EL LEGADO DEL PASADO
Tanto los estudios genéticos como la evidencia fósil acumulados durante esta década, nos proporcionan un retrato más diverso, más rico y más dinámico de nuestra propia especie. Desde su origen en África, hasta su hibridación con neandertales y denisovanos, Homo sapiens se retrata como un crisol de humanidades.
Es posible que muchas de las claves adaptativas de nuestro éxito en la conquista de un territorio más grande y un entorno más cambiante se encuentren, precisamente, en ese mestizaje cosmopolita que nos caracteriza desde hace al menos 200.000 años. Este mestizaje no solo no atentó contra nuestra identidad como especie, sino que es probablemente parte de su sello e idiosincrasia.
La evolución humana incide, precisamente, en que la diversidad, desde el punto de vista biológico, es una ventaja, un equipaje versátil del que la naturaleza puede tirar cuando las circunstancias requieren flexibilidad adaptativa. Las especies endogámicas y homogéneas son más propensas a mutaciones deletéreas y es posible, incluso, que el aislamiento prolongado de los neandertales en Europa, durante las edades de hielo, los haya hecho genéticamente más vulnerables.
Parte de la flexibilidad que hoy nos caracteriza nos fue transferida por otros humanos que hoy ya no existen. Somos presente y somos futuro, pero somos también herencia de quien ya no está.
A pesar de ser especies diferentes y probablemente reconocerse como tal, los humanos hibridaron con otras especies hoy extintas, tuvieron descendencia y cuidaron de ella. Es inevitable hacer una reflexión sobre la sociedad actual, particularmente ofuscada en trazar fronteras y marcar límites, entre individuos de la misma especie, que resultan más infranqueables que los que en principio dictaba la biología.
Con frecuencia, nuestra cultura y nuestras normas sociales avanzan hacia derroteros que parecen contradecir nuestro legado genético. ¿Cómo trataríamos a otra especie humana hoy en día? ¿Por qué somos la única especie superviviente? ¿Tendríamos sitio, en realidad, para otra humanidad distinta? ¿Espacio para la diferencia? ¿Cuál es nuestro nivel de tolerancia hacia la diversidad biológica y cultural?
Seguimos evolucionando. La selección natural sigue funcionando, pero hemos cambiado las presiones selectivas. Las presiones sociales tienen ahora un peso mayor que las ambientales, es más importante estar bien relacionado que disfrutar de una salud de hierro. Con el surgimiento de las técnicas de edición genética, el hombre disfruta ahora de un «superpoder» que todavía tiene que aprender a controlar.
Para ello la sociedad ha de plantearse un debate maduro y consensuado sobre hacia dónde queremos ir, pero ese debate necesita tener en cuenta nuestra propia historia evolutiva, comprender las peculiaridades y las claves del éxito de nuestra especie. Se mire por donde se mire, fue la diversidad y no la uniformidad la que nos hizo fuertes.
Más que nunca el ser humano lleva la batuta de su propio destino. Nos hemos convertido en nuestro propio genio de la lámpara. Podemos pedir ahora un deseo sobre nuestro futuro, pero es importante decidir qué queremos pedir. Presumimos de ser una especie inteligente, pero será lo que hagamos a partir de ahora lo que nos permita juzgar nuestra clarividencia. En diez, veinte, en cien años, nuestro pasado hablará de nosotros y será el pasado el que nos dé el verdadero veredicto sobre nuestra inteligencia.

Deja un comentario